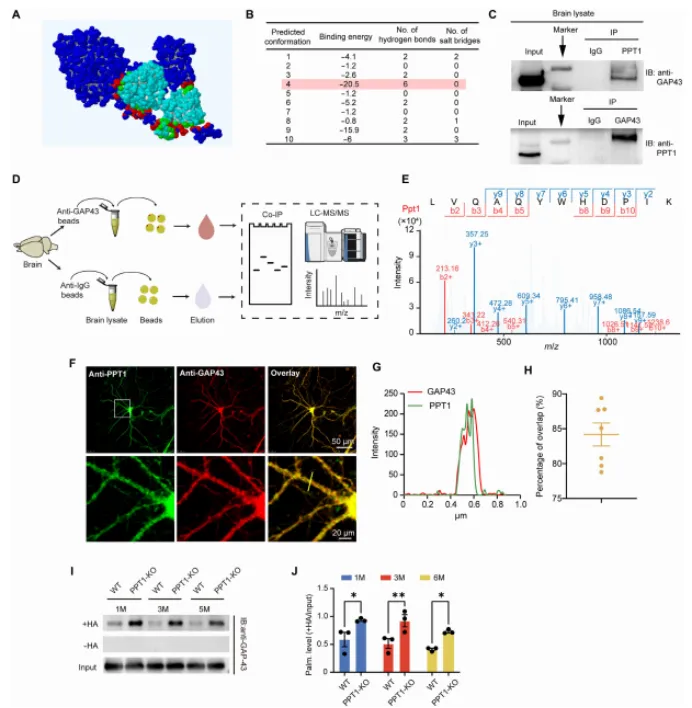
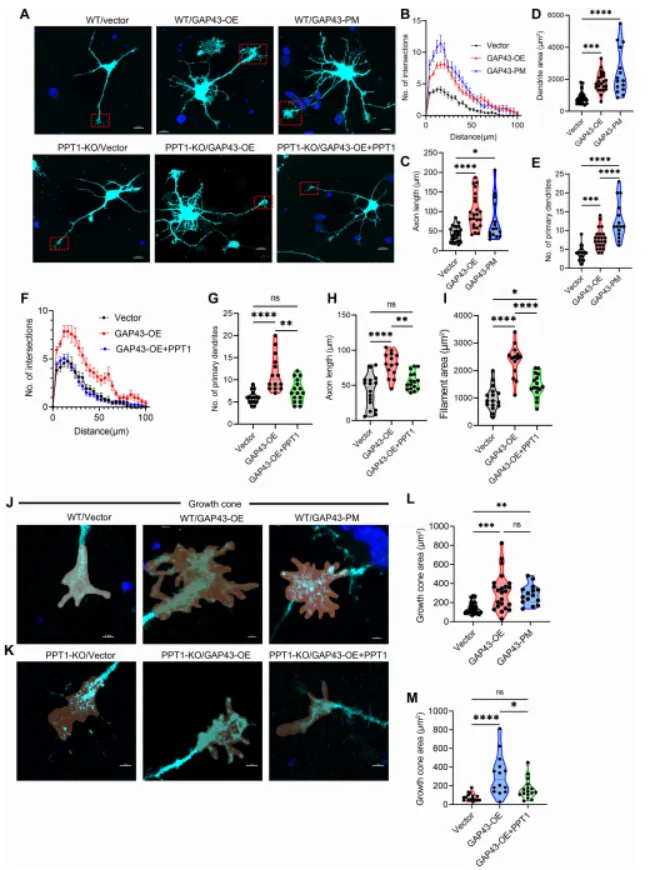
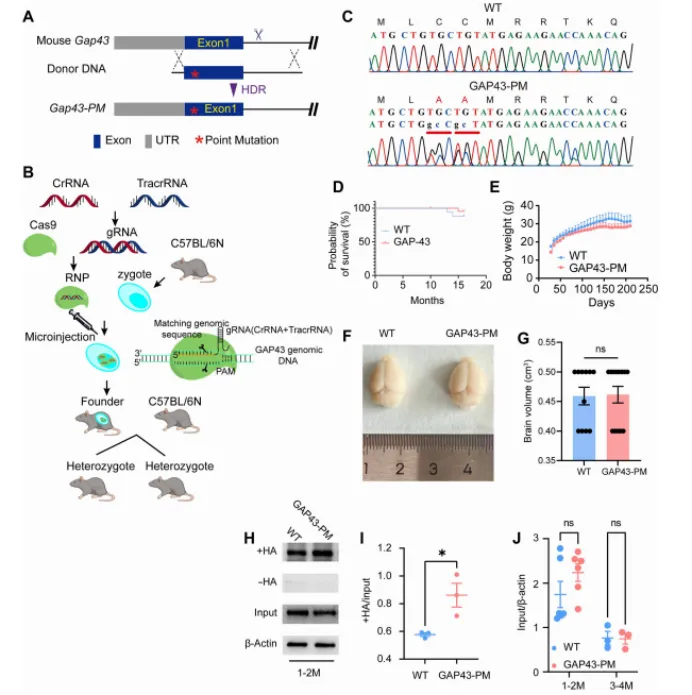
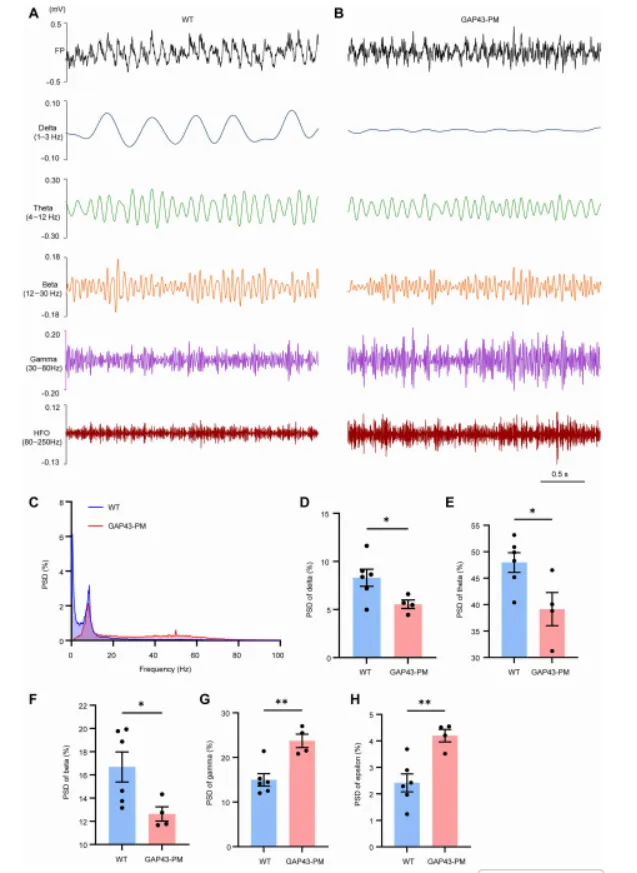
本研究通过结合生化实验、结构建模、利用 CRISPR-Cas9 构建的 GAP43 点突变小鼠(GAP43-PM)、PPT1 敲除小鼠(PPT1-KO)、电生理学以及行为学分析,作者证实 PPT1 可通过与 GAP43 的 Cys3 和 Cys4 位点相互作用介导其去棕榈酰化。
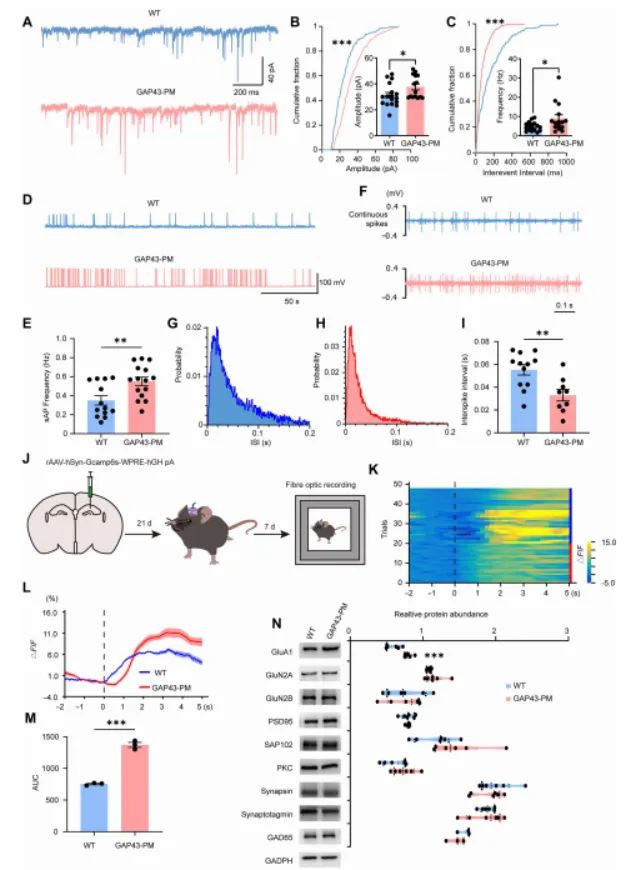
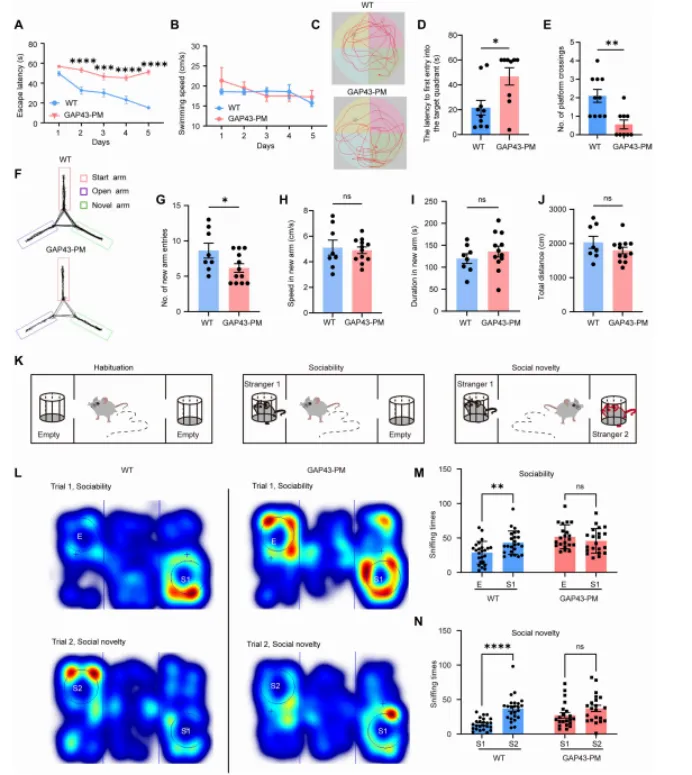
该相互作用的破坏会导致 GAP43 过度棕榈酰化(hyperpalmitoylation),进而引起过度的树突分枝(dendritic arborization)和异常生长锥扩张(growth cone expansion),增强谷氨酸能神经传递,并导致海马神经网络过度兴奋(hyperexcitability),最终造成认知功能缺陷,而这一过程并不伴随溶酶体贮积病理改变。此外,外源性 PPT1 能够减轻上述形态学和突触异常。本研究表明,PPT1–GAP43 去棕榈酰化通路对于维持神经环路稳态至关重要,其功能异常可能参与神经发育障碍的发生,并提示该通路可能成为棕榈酰化相关神经发育疾病的潜在治疗靶点。
这篇文章的主要亮点就在于用C3AC4A双点突变模拟“去棕榈酰化受阻”,而非直接敲除GAP43,从而将修饰状态与表达水平解耦。这一策略可广泛应用于我们翻译后修饰的相关研究: